| INTRODUCCIÓN
Ciertos autores han estudiado la emisión de veneno en las mordidas sucesivas a presas (Olrog, 1953; Naulleu y Detrait, 1976; Tun Pe et. al. ,1991; Hayes, 1992a). Algunos de estos estudios revelan que existe una disminución en la cantidad de veneno inoculado en las mordidas consecutivas ( Olrog, 1953; Tun Pe et. al. , 1991). El objetivo fundamental del presente trabajo se basa en el estudio de la emisión de ponzoña y las cantidades inyectadas en sucesivas presas, ratones, en sesiones de comportamiento alimentario de las especies autóctonas Bothrops alternatus ("Crucera") y Bothrops neuwiedii pubescens ("Yarará"). |
Docente Responsable: Melitta Meneghel - Sección Zoología Vertebrados Autores: Leandro Furest Nicolás Papa Mª Noel Sotelo Carolina Toranza
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fig. 1 Bothrops neuwiedi pubescens ("yarará") |
MATERIALES
Y MÉTODOS
Trabajamos con 3 ejemplares adultos de Bothrops neuwiedii pubescens (Fig. 1), con masas corporales: · N3 (775,5g) · N50 (1309g) · N93 (727,3g) y 2 ejemplares adultos de Bothrops
alternatus (Fig. 2), con masas corporales: · A91 ( 196,8) |
Fig. 2 Bothrops alternatus ("crucera") |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Todos los
ejemplares están habituados como mínimo a 5 años de cautiverio. Cabe
destacar que cada ejemplar está alojado en un habitáculo individual
con dimensiones (40x33x26) cm. Todos los ofidios tenían un ayuno de 9
días.
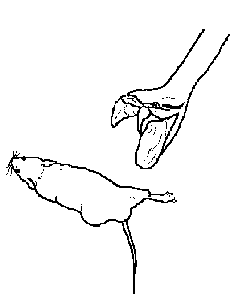
A cada ejemplar le fueron ofrecidos ratones blancos (Mus musculus) ad libitum. Las presas fueron pesadas en una balanza de triple brazo con una apreciación de 0,05g, antes y después de ser mordidos por el ofidio (Fig. 3), para hallar la masa de veneno inoculada. El tiempo transcurrido entre la mordida y la muerte de cada presa fue cronometrado. Una vez que el animal terminaba de deglutir y comenzaba la fase de búsqueda, se le ofrecía otra presa.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| RESULTADOS
Los tiempos de muerte, así como la masa de las presas, antes y después de la mordida se reseñan en la Tabla I y II. Por observación directa, constatamos que no todas las mordidas se produjeron en el mismo lugar sobre el cuerpo de la presa. En varios casos se verificó que la presa eliminó materia fecal, orina, o sufrió alguna pequeña pérdida de sangre.
|
Tabla I - Tiempos de muerte y masas de las presas ofrecidas a Bothrops neuwiedi pubescens en sucesivas mordidas. * La presa eliminó desechos, # Recibió dos mordidas.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Tabla II - Tiempos de muerte y masas de las presas ofrecidas a Bothrops alternatus en sucesivas mordidas. * La presa eliminó desechos.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fig. 3 Bote predatorio de una Bothrops (tomado de Sazima, 1989). |
DISCUSIÓN
En los casos en los que la presa mantuvo o disminuyó el peso registrado a priori, se pudo verificar que había mediado una pérdida de masa en forma de orina, materia fecal y, en algunos casos, sangre. Particularmente en el caso del tercer ratón ofrecido al ejemplar N93, asumimos que se produjo una punción intracardíaca, debido al lugar de la mordida y a la rapidez con la que perdió gran cantidad de sangre. Este hecho explicaría el corto tiempo de muerte. Debido a que la muestra de datos de la diferencia de masa de veneno fue pequeña (6 ratones en 17) (Tabla I y II), no pudimos concluir nada acerca de las cantidades efectivas de veneno emitidas. No podemos asumir que el veneno en las 2 horas y media, que duró la experiencia con cada ofidio conservó su potencia y concentración debido a lo observado en los ejemplares N50 y N93, ya que en ambos hubo un mayor aumento de masa en la presa acompañado de un aumento en el tiempo de muerte. Esto podría deberse asimismo a una menor eficiencia en las mordidas. Se constató que el segundo ratón ofrecido al ejemplar N3, fue mordido con un solo colmillo en el muslo, lo que prolongó el tiempo de muerte. Antes de que la presa muriera por el efecto de la ponzoña, el ofidio se acercó y la deglutió. El tiempo registrado en la Tabla I, no corresponde exactamente al de la muerte por envenenamiento. No registramos el peso post-inoculación del tercer ratón ofrecido al ofidio A23, debido a que perdió mucha sangre. Esta presa fue retenida brevemente durante el bote, tiempo en el que el ofidio pudo haber inoculado una mayor cantidad de veneno, lo que explicaría el corto tiempo de muerte ( Hayes, 1992b).
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Con el fin de
lograr el objetivo propuesto, fueron analizados para cada ofidio los
tiempos de muerte, de la serie de presas aceptadas por los mismos.
Excepto en el caso de la N93, no fue fácil determinar un patrón de tiempo que revelara un aumento progresivo del mismo en sucesivas mordidas (Hayes, 1992a). En cuanto al ofidio N3, a pesar de que el tiempo de muerte en la segunda presa se prolongó respecto a la primera, no se pudo concluir que se debiera a un descenso en el efecto del veneno, ya que esta mordida fue ineficiente (ver arriba). Respecto al ejemplar N50, se observó un aumento en el tiempo de muerte de las tres primeras presas. Cabe destacar que la tercer presa recibió una segunda mordida a los 76segundos de haber recibido el primer bote. En cuanto a la cuarta y quinta presa ofrecidas se constató una acentuada disminución en el tiempo de muerte; hecho que podría ser explicado por mordidas muy eficientes. Las presas ofrecidas a N93 demoraron sucesivamente más en morir, y fue en el caso en que más claramente pudimos verificar nuestra hipótesis. En la A23 se observó un patrón inverso al esperado, lo que podría explicarse por una mejora progresiva en la eficiencia de las mordidas. Entre las "Cruceras", la A91 nos permitió ver el comportamiento de emisión esperado. Los tiempos prolongados en la muerte de los ratones pueden deberse a la pequeña masa del ofidio, en comparación a los otros ejemplares.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
CONCLUSIONES
El tiempo de muerte aumenta en sucesivas mordidas a presas. Esto se constató en ambas especies. No todas las mordidas son efectuadas con igual eficiencia. La eficiencia de las mordidas puede variar en un mismo ofidio. La eficacia de la mordida pesa más en el tiempo de muerte que la cantidad de veneno inoculado (Hayes,1992a). Sería necesaria una muestra mayor, para poder analizar la cantidad de veneno inoculado con relación al tiempo de muerte.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BIBLIOGRAFÍA
Hayes,W. 1992a. Factors associated with the mass of venom expended by Praire rattlesnakes. (Crotalus v. viridis ) feeding on mice . Toxicon, 30 (4): 449-460. Hayes, W. 1992b. Prey-handling on envenomation strategies of Praire rattlesnakes (Crotalus v. viridis) feeding on mice and sparrows. Journal of Herpetology, 26 (4): 496-499. Naulleu, G. y Detrait, J. 1976. La fonction venimeusse chez Vipera aspis (L.) élevée en conditions experimentels artificielles. Bulletin de la Societé Zoologique de France, 101(4): 728. Orlog, C. 1953. Observações sobre a alimentação de uma cascavel ( Crotalus durissus terrificus Laur.) em cativeiro. Papéis Avulsos, 11(5): 41-44. Sazima, I. 1989. Comportamento alimentar da jararaca, Bothrops jararaca: encotros provocados na natureza. Ciência e Cultura, 41(5): 500-505. Tun Pe; Khing Aung Cho y Aye Aye Myint. 1991. Amount of venom injected by Russell´s viper (Vipera russelli) in killing prey. The snake, 23: 71-74. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||