COMPORTAMIENTO
ALIMENTARIO COMPARADO ENTRE COLUBRIDOS Y VIPERIDOS USANDO RATONES COMO PRESAS


INTRODUCCIÓN:
La conducta predatoria en serpientes varia según
a que familia pertenezca (NAULLEAU,1975).
Ante ratones como presas, los colúbridos utilizan
la constricción como principal forma de captura, mientras que los vipéridos
utilizan inyección de ponzoña. Estas conductas son consecuencia de la
evolución morfológica.
El comportamiento alimentario difiere con respecto
al tamaño de la presa, posición de captura, contacto inicial, etc. y
situaciones que se presenten en distintas circunstancias (Loop & Bailey,
1972; Diefenbach & Emslie, 1971).
Los sentidos captan diversas señales (químicas,
visuales, térmicas, táctiles) que son tomadas en cuenta en el momento de la
captura y reconocimiento de la presa.
Nuestro trabajo se basó en identificar y comparar
las distintas etapas del comportamiento alimentario de vipéridos y colúbridos.
|

|

|
|
Comienzo de la
ingestión |
Constricción e
inyección de ponzoña |
|
|
|
|
|
|

|
|
Ingestión |
Constricción |
|

|

|
|
Fin de
la ingestión |
Bostezo |
|

|

|
|
Ingestión |
Constricción
fuera del terrario |
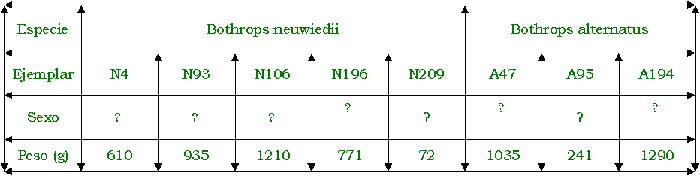
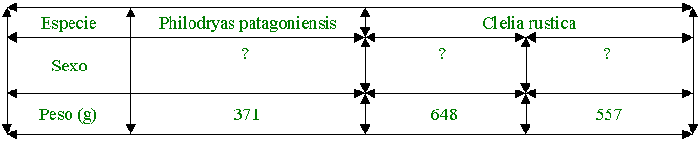
Se utilizaron tres individuos de Bothrops
alternatus, cinco individuos de Bothrops neuwiedii, una Philodryas
patagoniensis y dos individuos de Clelia rustica, de las cuales algunas fueron
colectadas en años anteriores y otras nacidas en cautiverio.
Las especies se encuentran alojadas en terrarios
de 41x33x26h con una base de cartón corrugado y agua a disposición en
recipientes de cerámica o plástico, a una temperatura aproximada de 27ºC.
Fueron alimentadas con dieciocho ratones blancos
con peso aproximado de 20g cada uno, puestos dentro de los terrarios mediante
una pinza de 30cm. Para medir el tiempo de las distintas etapas del
comportamiento alimentario se utilizó un cronómetro y los registros fueron
tomados con cámaras digitales y manual.
De dieciocho encuentros provocados, en uno de
ellos la serpiente se mostraba sumamente inquieta, no llegando a ingerir la
presa. En presencia de la misma comienza la alerta debido a los movimientos de
ésta.
Las diferencias entre las familias se inician en
la orientación, donde en vipéridos es más "cuidadosa" para
realizar un bote efectivo y lograr una mejor acción de la ponzoña debido a
que es su única forma de dar muerte a la presa. Sazima (1989) comprobó que
el 73% de los botes de Bothrops jararaca son en la región escapular. En colúbridos
los botes fueron sucesivos e imprecisos (llegando a morder su propio cuerpo)
porque los utilizan como forma de anclaje para constreñir y no tanto para la
inyección de ponzoña (ambas especies de culebras son opistoglifas).
Luego del bote los vipéridos sueltan la presa, se
retraen y entran en un estado de latencia a diferencia de los colúbridos que
generalmente constriñen de forma dextrógira o levógira. Observamos que dos
individuos alternaron su flanco de constricción, posiblemente debido a la
posición inicial de ataque. Esto es resultado de las diferencias en las técnicas
de captura y en la morfología de los ofidios (Willard, 1977).
El rastreo sólo se da en vipéridos porque los
colúbridos se mantienen en contacto con la presa.
Cuando los vipéridos encuentran la presa comienza
el reconocimiento, siendo éste más "minucioso" que en culebras
debido a que en la constricción éstas solo dejan una parte visible de la
presa.
La ingestión de los vipéridos fue sólo por
cabeza. Sin embargo en colúbridos observamos tanto por cabeza como por cola,
siendo el tiempo de ingestión por cola casi el doble que por cabeza
(Diefenbach & Emslie, 1971). Probablemente el tamaño relativo de las
presas con respecto a los colúbridos fuera pequeño (Loop & Bailey,
1972).
En Bothrops neuwiedii observamos luego del bote e
ingesta acomodamientos bucales, en Bothrops alternatus realiza además
bostezos y los colúbridos sólo presentan acomodamientos bucales luego de la
ingestión.
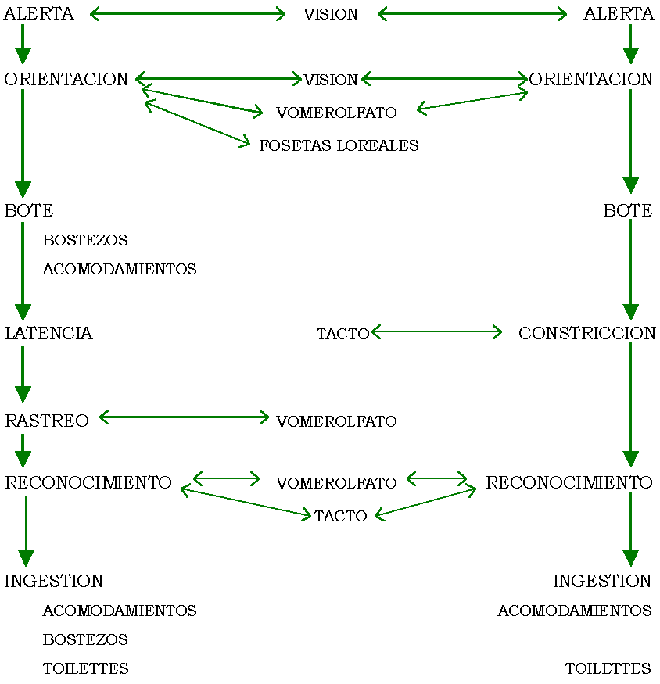
ETOGRAMA I:
Vipéridos
ETOGRAMA II: Colúbridos
Para ambas familias el único sentido involucrado
en la alerta es la visión.
El comportamiento "cuidadoso" de los vipéridos
en la orientación se debe a que dirigen la mordida hacia una determinada zona
del cuerpo de la presa.
La constricción dextrógira o levógira depende
del individuo y en la situación en la cual se encuentre.
El rastreo sólo se presenta en vipéridos.
En vipéridos generalmente la ingestión es por
cabeza porque puede reconocer el total de la presa a diferencia de los colúbridos
que sólo disponen de una parte de la presa.
En colúbridos la ingesta por cola requiere más
tiempo que por cabeza por el impedimento que presentan el pelaje y los
miembros.
Luego del bote o ingestión los vipéridos
presentan acomodamientos bucales o bostezos, los colúbridos presentan sólo
acomodamientos bucales.
BIBLIOGRAFÍA:
DIEFENBACH, C.O. & EMSLIE, S.G. 1971.
Cues influencing the direction of prey ingestion of the Japanese snake, Elaphe
climacophora (Colubridae, serpentes). Herpetologica, 27(4):461-466.
LOOP, M.S. & BAILEY, L.G. 1972. The
effect of relative prey size on the ingestion behavior of rodent-eating
snakes. Psychon. Sci., 28(3):167-169.
NAULLEAU, G. 1972. Comportément prédatéur
de la vipère à cornes (Aspis cerastes, Linné). Rev. Comp. Animal,
6:273-282.
NAULLEAU, G. 1975. La prédation chez les
serpents. Bulletin de la Société Zoologique de France, 100(4):682-683.
SAZIMA, I. 1989. Comportamento alimentar da
jararaca, Bothrops jararaca: encontros provocados na natureza. Ciência e
cultura, 41(5):500-505.
WILLARD, D.E. 1997. Constricting methods of
snakes. Copeia, 1977(2):379-382.

DOCENTE RESPONSABLE: Meneghel, Melitta - Sección
Zoología Vertebrados.